August 10, 2017
David M. Checkley, Rebecca G. Asch, and Ryan R. Rykaczewski
http://www.annualreviews.org/doi/10.1146/annurev-marine-122414-033819
Abstract
Anchovy and sardine populated productive ocean regions over hundreds of thousands of years under a naturally varying climate, and are now subject to climate change of equal or greater magnitude occurring over decades to centuries. We hypothesize that anchovy and sardine populations are limited in size by the supply of nitrogen from outside their habitats originating from upwelling, mixing, and rivers. Projections of the responses of anchovy and sardine to climate change rely on a range of model types and consideration of the effects of climate on lower trophic levels, the effects of fishing on higher trophic levels, and the traits of these two types of fish. Distribution, phenology, nutrient supply, plankton composition and production, habitat compression, fishing, and acclimation and adaptation may be affected by ocean warming, acidification, deoxygenation, and altered hydrology. Observations of populations and evaluation of model skill are essential to resolve the effects of climate change on these fish.
INTRODUCTION
Climate change and fishing are the two dominant processes by which humans affect small pelagic fish. Both climate change and fishing have increased since about 1850. Industrial fishing and greenhouse gas (GHG) emissions rose together as technology developed and both fish and fuel were abundant and accessible. However, although fishing can ideally result in production equilibrium—i.e., removals of fishes are compensated for by increased fish production at lower fish densities—increasing GHG concentrations result in climate disequilibrium. For the coming generations, although capture fisheries are unlikely to increase much beyond current levels, the earth will warm, the oceans will acidify, and hydrology will change. How will small pelagic fish respond?
Anchovy and sardine made up 52% of landings of small pelagic fish and 13% of global capture fisheries in 2012 (FAO 2014). The bycatch of anchovy and sardine fisheries is relatively low and their efficiency is relatively high (Kelleher 2005), with relatively low GHG emissions per landed weight (Parker & Tyedmers 2015), both of which result from the schooling behavior of the fishes and the purse seine fisheries that target them. They are used for aquaculture feed, industrial oil, health supplements, and human consumption (Barange et al. 2014). Anchovy and sardine are key forage for marine predators, including fish, squid, marine mammals, and seabirds. They are of increasing interest for conservation because of their perceived role as critical forage for charismatic megafauna (e.g., marine mammals and seabirds) in many regions, and are often in the public eye (Cury et al. 2011). Here, we consider anchovy and sardine species that complete all or most of their life cycle in water of oceanic salinity. The areas occupied by these species include all oceans and several inland seas and, collectively, these species constitute a large majority of the biomass of all anchovy and sardine.
Understanding the evolution and past variability of anchovy and sardine and their causes, including climate and fishing, is necessary to detect and predict the responses of these fishes to atmospheric and oceanic changes caused by GHG emissions. These changes include the warming of the earth; changes in ocean biogeochemistry, including acidification and deoxygenation; and alteration of the hydrological cycle.
This review consists of three parts. The first part addresses the past, including anchovy and sardine evolution, their populations, and fishing. The second part addresses the present relations of anchovy and sardine to the environment. Finally, the third part addresses the future, including projected effects of climate change on anchovy and sardine, observations to date consistent with these projections, and recommended actions.
THE PAST
Anchovy and sardine evolved over millions of years (Grant & Bowen 1998), during which the earth’s climate varied naturally. GHG emissions have now accelerated changes not only in temperature but also in ocean chemistry (Gattuso et al. 2015) and hydrology (IPCC 2013). The past evolution and fluctuations of anchovy and sardine in a naturally varying ocean may inform us about their potential to respond to future rapid change (Munday et al. 2013).
The Evolution of Anchovy and Sardine
Anchovies (Engraulis) and sardines (Sardinops, Sardina, and Sardinella) have been described as having “shallow population histories in deep evolutionary lineages” (Grant & Bowen 1998). This implies that the two taxa and their species evolved millions of years ago but have relatively recently, and in places repeatedly, established populations. Molecular genetics indicate that anchovy and sardine speciated tens of millions of years ago (Grant & Bowen 1998).
Sardinops and Sardina split from a common ancestor approximately 19 million years ago (Bowen & Grant 1997). Approximately 500,000 years ago, Sardinops colonized five main locations: off California, Japan, Chile-Peru, Australia, and South Africa (Bowen & Grant 1997). Sardina is the European sardine; Sardinella includes tropical and some subtropical sardines (Parrish et al. 1989).
Engraulis separated from Encrasicholina in the Americas tens of millions of years ago (Grant et al. 2005). Engraulis split into Old World and New World groups approximately 3–8 million years ago (Grant & Bowen 2006). Old World (Japan, Australia, South Africa, and the Northeast Atlantic) and New World (California, Chile-Peru, and Argentina-Brazil) populations were established hundreds of thousands of years ago (Grant & Bowen 1998).
Several traits common to both anchovy and sardine are noteworthy. Most species are r selected—i.e., they have a large reproductive potential and thus are capable of rapid population growth (Murphy 1968). Most species are fecund, with serial spawning of many pelagic eggs (Blaxter & Hunter 1982). High fecundity allows for high recruitment and rapid population growth under conditions favorable for survival in a variable environment (Murphy 1968). Anchovy are smaller, shorter lived, and less fecund than sardine (Lindegren et al. 2013, Murphy 1968). Anchovy and sardine have similar life histories, composed of a brief pelagic egg stage, hatching, yolk-sac larvae, feeding larvae, metamorphosis, juveniles, and mature adults. Both anchovy and sardine have elongated and compressed bodies. Both are silvered, with species-specific, guanine-containing scales whose accumulation in anoxic sediments provides a record of past abundance (Field et al. 2009). Both taxa school, particularly in daytime, to minimize predation (Kaltenberg & Benoit-Bird 2009). However, they also differ in key traits.
Two noteworthy divergent traits are feeding and migration. Anchovy and sardine eat phytoplankton and zooplankton at or near the base of the food web by filtering particles from water, biting individual particles, or both (van der Lingen et al. 2009). Gill rakers are used for filtering. Anchovy have coarse filters, capable of retaining relatively large particles; sardine have fine filters, capable of retaining smaller particles. Anchovy both filter and bite, whereas sardine primarily filter. Consequently, sardine consume smaller plankton than anchovy (Rykaczewski & Checkley 2008, van der Lingen et al. 2009). Sardine are generally larger and faster swimmers than anchovy (Blaxter & Hunter 1982). Consistent with this, sardine often migrate farther than anchovy (Checkley et al. 2009b). This movement may consist of a return migration following passive transport from a spawning location during the planktonic egg and larva stages [e.g., the Japanese sardine migration between spawning habitats off Honshu and the juvenile habitat in the Kuroshio-Oyashio Extension (Watanabe et al. 1995)] or a migration between feeding and spawning grounds by older fish [e.g., the Pacific sardine migration between California and Washington or British Columbia (Demer et al. 2012)]. The sardine run along the east coast of South Africa is a migration noteworthy for its regularity and human interest (van der Lingen et al. 2010). By contrast, anchovy are smaller and migrate less (e.g., from onshore to offshore). The body-size differences are also consistent with the differing bioenergetic requirements of their primary feeding modes (van der Lingen et al. 2009). The gliding associated with filter feeding confers a bioenergetic advantage to the larger sardine, whereas the picking motion is bioenergetically advantageous for the anchovy.
Populations
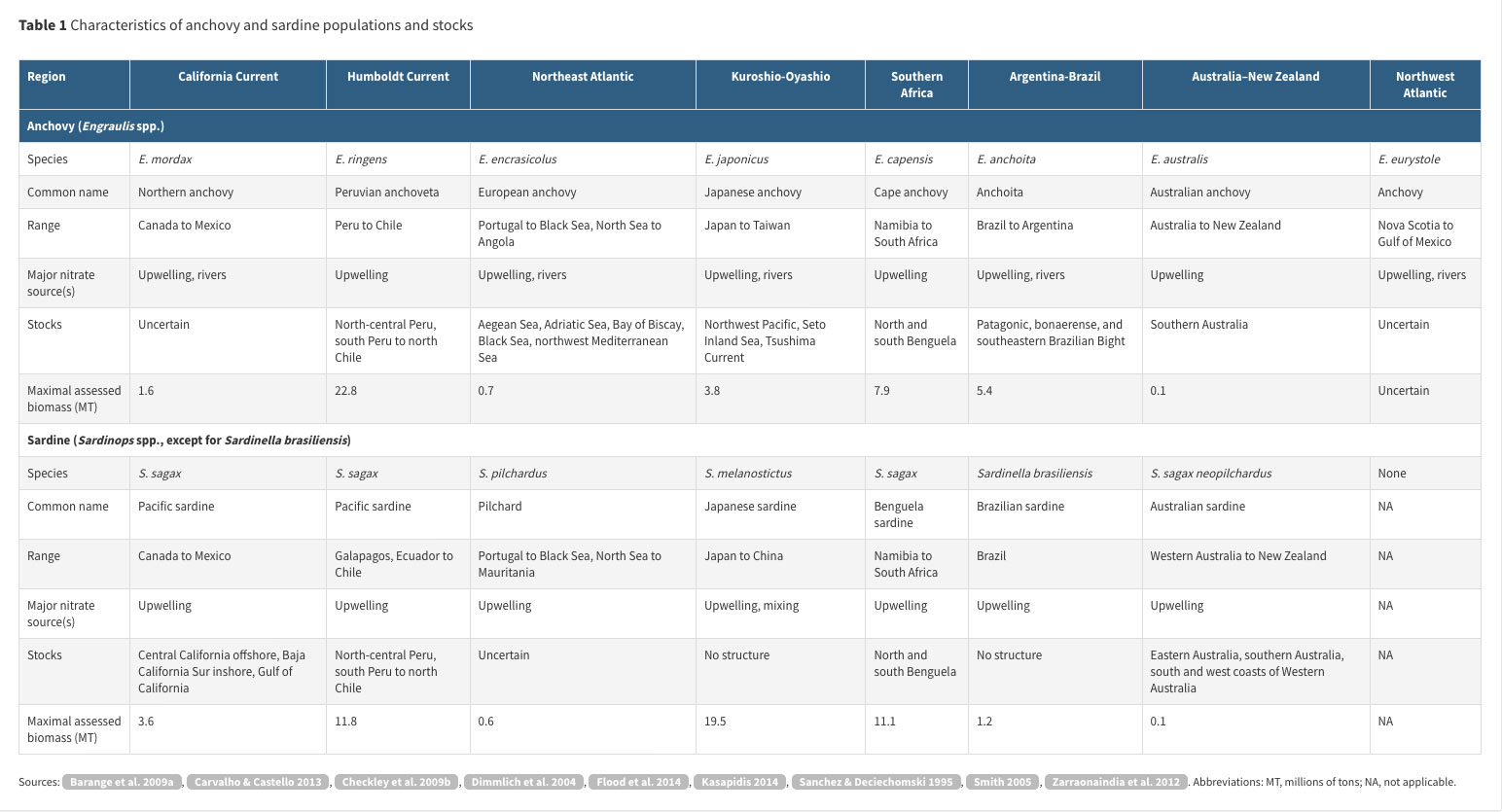
Both anchovy and sardine populations exist in most temperate and productive coastal regions in the oceans and major seas (Figure 1, Table 1). A notable exception is the absence of sardine from the Northwest Atlantic. Within populations, stocks (Kasapidis 2014, Parrish et al. 1989) are usually the target of fisheries.
View: Table 1
{kind=link}
Characteristics of anchovy and sardine populations and stocks
Anchovy populations in all regions have multiple stocks (Kasapidis 2014, Zarraonaindia et al. 2012). The world’s largest single-species fishery, with a maximal biomass of 22.8 million tons (MT), is for Peruvian anchoveta (Table 1). Among anchovy and sardine stocks, those with the fifth-, sixth-, and seventh-ranked maximal estimated biomass levels caught by fisheries are the South African anchovy (7.9 MT), Argentine-Brazilian anchoita (5.4 MT), and Japanese anchovy (3.8 MT), respectively. Most anchovy stocks are coastal (i.e., in areas of the ocean bounded by land on one side).
Sardine populations have varying degrees of stock structure within regions (Kasapidis 2014, Parrish et al. 1989, Smith 2005). Among anchovy and sardine stocks, the Japanese sardine has the second-largest maximal biomass caught by fisheries (19.5 MT). The third-, fourth-, and eighth-largest maximal biomasses are for sardines off Peru (11.8 MT) and Namibia (11.1 MT) and in the California Current (3.6 MT), respectively.
Each region with large populations of anchovy and sardine is nourished by upwelling, the upward movement of nutrient-rich water from beneath the mixed layer into the euphotic zone, often stimulated by winds. However, it is notable that the Japanese sardine, which is second in maximal landings only to the Peruvian anchoveta, inhabits the Kuroshio-Oyashio Transition Area, which undergoes deep, convective mixing each winter (Yasuda 2003). Many anchovy populations occur along coasts with rivers, notably the northern, European, and Japanese anchovies and the Argentine-Brazilian anchoita. The greater population structure of anchovy compared with sardine may be related to their greater affinity for coastal waters and (in some cases) rivers compared with sardine, which often occur farther offshore (Rykaczewski & Checkley 2008, Swartzman et al. 2008). The offshore distribution of sardine, which have finer gill rakers, is consistent with plankton communities that are composed of relatively small individuals in offshore waters (Rykaczewski & Checkley 2008, Zarauz et al. 2009). The larger size and longer migrations of sardine may also contribute to the lower genetic heterogeneity within its populations.
The current populations of anchovy and sardine were established within the last 500,000 years and show low genetic diversity (Bowen & Grant 1997). The “shallow life history” of most anchovy and sardine populations indicates that they fluctuated greatly in size and, for some populations, were extirpated and recolonized (Grant & Bowen 1998). Climate variability, particularly Milankovitch cycles of the past 500,000 years and the associated variations in ocean temperature and sea level, has been proposed to be an important factor in the past ebb and flow of populations of Old World anchovies (Grant & Bowen 2006). Warming and cooling shifted the suitable habitats, and a low sea level restricted movement of populations. Refugia are likely important for population survival at a small size (Lecomte et al. 2004). Examples of refugia for current populations include the Galapagos for the Peruvian anchoveta (Bertrand et al. 2004), the Pacific coast of Baja California for the Pacific sardine (Lluch-Belda et al. 2003), and river plumes for the European sardine (Bellier et al. 2007).
The cape anchovy off South Africa is noteworthy for the lack of a refugium (Grant & Bowen 2006). The cape anchovy consists of two genetic types, each of which is more closely related to the European anchovy than to the other cape anchovy type. The cape anchovy appears to have been repeatedly extirpated and refounded, the source being Europe. An explanation may be that the cape anchovy habitat in the Benguela Current System is bounded by two warm currents: the Angola Current to the north and the Agulhas Current to the east. Under warming, the cape anchovy is unable to shift poleward, lacking a refugium. The cape anchovy off South Africa may thus have been extirpated during past warm periods and be unusually vulnerable to ocean warming from future climate change.
Fluctuations Over Time: The Paleontological Record
Coastal basins with high primary productivity and low exchange of bottom water often contain anoxic sediments in which fish parts are preserved, forming a record of past fish occurrence (Soutar & Isaacs 1969). Sediment cores reveal annual varves that contain fish scales, bones, and otoliths (Field et al. 2009, Finney et al. 2010, Jones 2016, Soutar & Isaacs 1969). Time series of the deposition rate of fish scales and otoliths have been constructed for periods of centuries to millennia (Field et al. 2009, Jones 2016). These can be used to characterize fluctuations in populations of small pelagic fish prior to industrialized fishing and GHG emissions. Some anoxic basins are at or near regions of high production and the known, modern-day spawning locations of anchovy and sardine, allowing comparisons of deposition rates and populations (Field et al. 2009). Anchovy and sardine populations expand and contract laterally as they grow and shrink, respectively (Barange et al. 2009b, MacCall 1990). The degree to which sedimentation of fish parts represents changes in population size or distribution is uncertain. In general, population size varies with scale deposition rate off California and Peru (Field et al. 2009). Although not exact, the deposition rate of fish scales is often used as a proxy for the abundance of anchovy and sardine over the past few millennia.
The Santa Barbara Basin is adjacent to the dominant spawning habitat of anchovy and sardine on the US West Coast. The basin flushes infrequently and is typically anoxic (Goericke et al. 2015). The deposition rates of anchovy and sardine scales have fluctuated over the past two millennia with weak but significant positive correlation (Baumgartner et al. 1992). Periods have existed when both rates are high, both are low, or one is high and the other is low. Modes of variation with a 50–80-year period exist for both species, similar to that of the North Pacific climate (Minobe 1997).
Analysis of fish scales in sediment cores from other locations also indicates that anchovy and sardine fluctuated prior to climate change and fishing. Anchovy and sardine abundance have alternated in the Gulf of California since at least 1,730 years before present (YBP) (Holmgrenurba & Baumgartner 1993). Anchovy and, to a lesser degree, sardine scales in the sediments of basins off Peru and Chile show variations prior to fishing on a range of scales over the past 1,500 years (Guiñez et al. 2014, Valdes et al. 2008). A core to approximately 600 YBP from Walvis Bay, Namibia, was dominated by sardine scales that varied with an approximate period of 25 years (Shackleton 1987).
In summary, anchovy and sardine scales accumulated at sites off California, Peru, Chile, and Namibia at variable rates both before and after the start of human fishing activity. This variability is likely the response of the anchovy and sardine populations to climate variation (Baumgartner et al. 1992, Lindegren et al. 2013).
Fishing Peaks and Greenhouse Gases Accumulate
The period since about 1950 is one of accelerating GHG accumulations and asymptotic landings by capture fisheries (Figure 2). Rates of emission and accumulation of GHGs in the atmosphere and ocean continue to increase (IPCC 2013). pCO2 in the atmosphere has increased from a preindustrial concentration of 280 ppm to more than 400 ppm (Keeling 2016). Seawater pH has decreased by 0.11 units, equivalent to a 30% increase in the concentration of hydrogen ions (IPCC 2013). Simultaneously, the annual global fisheries catch reached a maximum of 130 MT in the mid-1990s and has since fluctuated around 110 MT (Pauly & Zeller 2016). All major stocks of anchovy and sardine have been fully exploited (Herrick et al. 2009). The catch of individual stocks of anchovy and sardine has fluctuated in a way that is consistent with fluctuations estimated from scale deposition rates in the geological record, with Namibia a notable exception. The fisheries off Japan, the west coast of North America, Peru, South Africa, and Northwest Africa have varied on a scale of 50–80 years, with decline followed by recovery (Barange et al. 2009a). However, the sardine fishery off Namibia, which peaked in 1993, has not recovered (Boyer & Hampton 2001, Kirchner et al. 2009).
CURRENT UNDERSTANDING
Fisheries have led to observations, analysis, and modeling as means to understand the spatial and temporal variation of populations of small pelagic fish. Habitats have been described and mechanisms for population variability proposed (Checkley et al. 2009b, MacCall 2009). The resultant understanding can be used to create models to predict future states and thus inform management and policy decisions under climate variability and change. This understanding also shows how fisheries management has a limited ability to control fluctuations in stock size, consistent with the primary control of those stocks being from the bottom up (i.e., climate) rather than from the top down (i.e., fishing).
Habitats
Anchovy and sardine require plankton for food. A useful way to classify populations is by the source of new nitrogen on which their food relies (Figure 3). New production is based on nitrogen from outside the system (Dugdale & Goering 1967). Fishing removes nitrogen in anchovy and sardine from their habitat. At a steady state, an upper limit on fish production is based on the input of new nitrogen (Ryther 1969). We hypothesize that variation in the supply of new nitrogen among habitats influences the magnitude of primary production, its temporal variability, the composition of the plankton, and ultimately the productivity of anchovy and sardine populations. These sources are projected to vary with climate change because of changes in winds, stratification, river flow, and the nutrient content of the source water.
Anchovy are present where new nitrogen is supplied primarily by coastal, wind-driven upwelling or rivers (Figure 1, Table 1). Examples where coastal, wind-driven upwelling is the dominant source in anchovy habitats include Peru, Japan, the California Current System, Europe, South Africa, and southern Australia. The anchoita is noteworthy for its close association with the Río de la Plata (de Macedo-Soares et al. 2014). Many European anchovy stocks have river associations, e.g., in the Bay of Biscay (Bellier et al. 2007), the northwest Mediterranean Sea (Agostini & Bakun 2002), and the Aegean Sea (Lloret et al. 2004). The northern anchovy habitat includes water affected by the plume of the Columbia River (Emmett et al. 2005). Anchovy spawning and migration off China and Taiwan are related to freshwater (Tsai et al. 1997, Tu et al. 2012). Anchovy eggs are found in salinities as low as 15 off Japan (Oozeki et al. 2007). Thus, although upwelling is the principal source of new nitrogen in the anchovy habitats, rivers also contribute.
Sardine generally occur farther offshore than anchovy (Gutierrez et al. 2007, Rykaczewski & Checkley 2008). New nitrogen is supplied in these habitats by upwelling or deep mixing. The Pacific sardine in the California Current System inhabits a region dominated by wind-stress-curl-driven upwelling, in which the offshore increase in wind speed causes slow but widespread upwelling (Rykaczewski & Checkley 2008). This process has not been quantified in other regions but may be common, given the ubiquity of wind-stress-curl-driven upwelling (Chelton et al. 2004). The Japanese sardine is noteworthy for its occurrence in the Kuroshio-Oyashio Transition Area (Watanabe 2009). New nitrogen is supplied in this region by deep convective mixing in winter and lateral advection (Yasuda 2003). The depth of mixing is positively related to sardine survival (Nishikawa & Yasuda 2008).
Each region has a dominant source of new nitrogen that is reflected in the stock of anchovy or sardine that has the largest maximal abundance (Figure 1, Table 1). In the Humboldt Current region, the Peruvian anchoveta habitat is dominated by strong wind-driven upwelling owing to its proximity to the equator, coast orientation, and persistent winds (maximal biomass 22.8 MT). In the Northwest Pacific, the Japanese sardine, which has a large population size, inhabits the Kuroshio-Oyashio Transition Area, where new nitrogen is supplied by deep convective mixing and lateral advection (maximal biomass 11.8 MT). In the Benguela Current region, the Namibian sardine habitat is dominated by wind-driven upwelling (maximal biomass 11.1 MT). The Argentine-Brazilian anchoita is present in a region where both the Río de la Plata and upwelling are sources of new nitrogen (maximal biomass 5.4 MT). The Pacific sardine is dominant in the California Current region, where wind-stress-curl-driven upwelling is the primary source of new nitrogen (3.6 MT). Hence, either anchovy or sardine has shown maximal biomass in the period of fisheries in each region, each with a distinct source of new nitrogen.
How will upwelling, mixing, and rivers, as sources of new nitrogen for anchovy and sardine, respond to climate change? Anchovy are more closely associated with land, including coasts, capes, orographic features, and rivers, all of which are geographically fixed. Sardine rely more on open-ocean processes, away from land. Hence, anchovy have different constraints than sardine in their ability to respond to latitudinal shifts in water properties (such as warming) and hydrology. It is necessary to project air-sea fluxes of temperature and momentum (wind), which influence upwelling and mixing, and river discharge, as well as the nutrient content of the source water, to project new production in anchovy and sardine habitats and hence their potential productivity.
Mechanisms of Fluctuations
Mechanisms causing fluctuations of anchovy and sardine worldwide have received much attention (Alheit & Bakun 2010, Alheit et al. 2009, MacCall 2009). Although these are not the primary focus of this review, they are useful to create a conceptual understanding, which can then inform future projections of the responses of anchovy and sardine to climate change. Mechanisms vary in their specificity (e.g., species, populations, and stocks) and processes. A general theme is the relative importance of bottom-up and top-down control. A prevailing view is that the fluctuations of the size of anchovy and sardine stocks have been controlled primarily by climate variability (bottom up) and secondarily by mortality (top down) and intrinsic traits of the species. This view is consistent with both the paleontological record and observations during the period of fishing. The paleontological record indicates that, given a variable climate, including temperature, wind, and hydrology, fluctuations of anchovy and sardine stocks are inevitable. However, the characteristics of these fluctuations are likely affected by fishing and, at times, other sources of mortality. In particular, the recovery of stocks from low to high abundance may be slowed or prevented by even modest mortality caused by directed fishing, bycatch, or other factors, including marine mammals, seabirds, jellyfish, and disease (Boyer et al. 2001, Essington et al. 2015, Lindegren et al. 2013, MacCall et al. 2016, Ward et al. 2001). Similarly, the rate of decline of a stock caused principally by changes in climate may be accelerated by mortality of the types just listed (Essington et al. 2015, Lindegren et al. 2013, Pinsky & Byler 2015). Finally, the duration of a period of low abundance may be prolonged by enhanced mortality (Essington et al. 2015).
Fluctuations of anchovy, sardine, the environment, and their relationships have been described and analyzed, yet mechanistic understanding is incomplete. Ocean temperature has been shown to be important to explain fluctuations in anchovy and sardine in the North Pacific (Chavez et al. 2003, Jacobson et al. 2005, Lindegren et al. 2013, Takasuka et al. 2007), the Humboldt Current System (Alheit & Niquen 2004), and the Black Sea (Guraslan et al. 2014). Pacific sardine production was more closely related to upwelling caused by wind-stress curl than to coastal wind stress (Rykaczewski & Checkley 2008). Sardine recruitment off Namibia was related to ocean temperature and wind (Daskalov et al. 2003) and the area of suitable habitat (Kirchner et al. 2009). Proxies for the Atlantic meridional overturning circulation and the associated nutrient supply to surface waters were related to the catches of major anchovy and sardine fisheries in the Atlantic and Pacific (Kamykowski 2012). Zooplankton displacement volume explained variation in anchoveta population size off Peru for 1961–2005 (Ayon et al. 2004). Cumulatively, these studies indicate the importance of temperature and food to populations of anchovy and sardine worldwide and thus control from the bottom up by climate.
Top-down effects comprise mortality from natural predators and fishing. Anchovy cannibalize their eggs and larvae (Takasuka et al. 2004). Intraguild predation has been proposed to contribute to the alternation of abundances of anchovy and sardine (Irigoien & de Roos 2011). Juvenile albacore have been estimated to consume up to 17% of northern anchovy prerecruitment biomass in the California Current System (Glaser 2011). Boobies, cormorants, and pelicans were estimated to consume 14% of anchoveta production before the fishery and 2% during the fishery in the Humboldt Current System (Jahncke et al. 2004). Australian salmon were estimated to maximally consume 15% of Australian sardine (Hughes et al. 2014). Jellyfish have been proposed to both prey on the young stages and compete for the food of anchovy and sardine (Lynam et al. 2006, Roux et al. 2013). Sharks, seabirds, and marine mammals feed on sardine during the sardine run off South Africa (O’Donoghue et al. 2010). Fishing on top predators and anchovy affected the food web of the Black Sea by reducing their predation on the plankton, allowing blooms of competing gelatinous zooplankton (Daskalov et al. 2007).
Direct competition for food between anchovy and sardine appears to be rare. Anchovy and sardine have different feeding selectivities. Other taxa—particularly vertically migrating midwater fish and, at times, jellyfish—are as important (if not more important) competitors (Koslow et al. 2014, Robinson et al. 2014). Also, populations of anchovy and sardine are, in general, spatially and temporally separate (de Macedo-Soares et al. 2014, Gutierrez et al. 2007, Rykaczewski & Checkley 2008, Swartzman et al. 2008).
Anchovy and sardine populations have been characterized as wasp-waist, exerting pronounced effects on both their planktonic prey and predator populations (Cury et al. 2000). The top-down effect may exist at small but not large scales (Atkinson et al. 2014, Ayon et al. 2008). Moreover, increasing estimates of the abundance of midwater fish that migrate daily to the surface to feed on the plankton indicate that anchovy and sardine may not control the zooplankton, as implied by wasp-waist dynamics (Koslow et al. 2014).
Most hypotheses about anchovy and sardine population dynamics have been built around correlations without strong mechanistic understanding. This might be sufficient if future climate change were to be similar to past climate variability and if we had a sufficient duration of high-quality historical observations to represent all modes of natural variability. Neither is likely true. We still lack a complete understanding of responses to natural variability, especially at low frequency. Future climate modes may differ from those of the recent historical period, as has occurred over centennial scales (MacDonald & Case 2005). Hence, we need a mechanistic understanding that builds on the growing knowledge of biogeochemistry, plankton composition and trophic interactions, fish morphology, and bioenergetics as well as our best estimation of future conditions. Multiple mechanisms for population fluctuations may exist, and an understanding built on correlations between landings and the environment is unlikely to suffice.
Models
Models are necessary for prediction, which in turn is needed to inform decisions under climate change. We briefly consider two types of models: statistical and mechanistic. Both are useful, albeit for different purposes and at different spatial and temporal scales. Models must be at a spatial scale consistent with those of fish populations and stocks and their environment.
Statistical models normally assume that the relationships between model parameters and the response variable do not vary over time. Thus, statistical relationships derived from past observations are used to create a model that explains past variability and allows for near-term forecasts. The properties of the system and knowledge of initial conditions limit the degree of forecasting in regard to time and detail. These two limitations make statistical models most useful for near-term predictions.
Two examples are stock-recruit relationships and equation-free models. The relationship between the biomass of a fish stock and the number of fish that recruit to its fishery at a certain size or age is fundamental to fisheries management (Quinn & Deriso 1999). This density-dependent relationship assumes that the number of progeny that recruit to a fishery is proportional to the number of eggs spawned until an environmental carrying capacity is reached. Departures from this relationship are density independent and attributed to the environment. Implicitly, the stock-recruit relationship does not account for climate variability and change, although environmental terms can be added to expand this type of model. Equation-free models reconstruct the behavior of systems directly from time-series data without the standard assumption of equations for the relationships between variables and have been applied to the Pacific sardine (Deyle et al. 2013). However, such models are unable to project the future state of ecosystems on the scale of climate change.
Mechanistic models are numerical representations of the dynamical processes within a system using equations to specify relationships between variables. Such models vary greatly in complexity, ranging from those with few parameters (e.g., the Lotka-Volterra model) to those with many parameters (e.g., end-to-end models). Of interest are models capable of representing the effects of climate variability and change on populations and stocks of anchovy and sardine. We briefly review models of anchovy and sardine that are not time dependent (i.e., equilibrium models) and those that depend on time. We acknowledge that this is but one classification of models.
Equilibrium models, including Ecopath (Pauly et al. 2000) and related types, have been used to characterize food webs with anchovy and/or sardine across many systems. Ecopath has been adapted to include bottom-up forcing (Steele & Ruzicka 2011) and time dependence (Ecosim; Pauly et al. 2000). A general use of this model has been to evaluate the effects of changes to the ecosystem caused by fishing. This type of model has shown that the Benguela Current System has undergone major reorganizations, particularly off Namibia from the 1960s (abundant sardine) to the 2000s (few sardine, increased gobies and jellyfish) (Shannon et al. 2009). The Atlantis end-to-end model has been applied to the southern Benguela Current System (Smith et al. 2015). Similar models have been used to characterize the food web off southern Australia, which includes a major sardine stock, with particular attention to interactions between species, including seals and squid (Goldsworthy et al. 2013). The Mediterranean Sea has been fished heavily, and equilibrium models provide insight into the importance of top-down effects on the ecosystems, including on anchovy (Halouani et al. 2015, Piroddi et al. 2015). The northern California Current System was modeled in detail using both fisheries-dependent and fisheries-independent data and including both climate (sea-surface temperature) and fishing, showing the overriding importance of bottom-up processes (climate) (Field et al. 2006) and jellyfish (Ruzicka et al. 2012). Ecopath-type models are particularly useful for comparing systems (Shannon et al. 2009). Collectively, these studies are consistent with climate regulation of the bottom-up control of anchovy and sardine and their ecosystems but also highlight how they are affected by fishing and other forms of predation.
Many dynamical models exist for systems with anchovy and sardine. There has been a progression from less complex, yet insightful, one-dimensional models (Oguz et al. 2008) to more complex models capable of recreating important characteristics of anchovy and sardine dynamics (Fiechter et al. 2015, Rose et al. 2015). The main elements characterize the water flow and properties in time and space, usually using regional ocean models; the basic biogeochemistry, using nutrient-phytoplankton-
Such models exist for all regions. The Black Sea was invaded by the ctenophore Mnemiopsis leidyi, and the anchovy there declined but subsequently began to recover. These dynamics, based on a model, were attributed to competition, predation, and habitat changes (Oguz et al. 2008). A model described European anchovy recruitment in the northwest Mediterranean Sea as depending on the place of spawning and drift of eggs and larvae as well as on the Ebro River (Ospina-Alvarez et al. 2015). A full life-cycle model of European anchovy in the northern Aegean Sea successfully reproduced observed characteristics of the stock, including migration (Politikos et al. 2015). Retention and habitat, including future pCO2, have been indicated to be important in the Benguela Current System (Brochier et al. 2013, Travers-Trolet et al. 2014). The effect of El Niño on the anchoveta in the Humboldt Current System has been modeled (Xu et al. 2015). The spawning location off Japan and temperature and food in the Kuroshio Extension and Kuroshio-Oyashio Transition Area were shown to be important to the survival and growth of sardine larvae and juveniles (Nishikawa & Yasuda 2011, Okunishi et al. 2012). Climate effects on the northern anchovy and Pacific sardine in the California Current System were modeled by combining general circulation, regional ocean, individual-based, bioenergetic, and fishing models (Fiechter et al. 2015, Rose et al. 2015). The past dynamics of anchovy and sardine, including long-term fluctuations, were successfully reproduced. The success of this model depends in part on knowledge and representation of biological traits distinguishing anchovy and sardine. A simpler model of the same populations recreated both paleontological (before fishing) and recent (with fishing) fluctuations of anchovy and sardine (Lindegren et al. 2013).
Collectively, these and other models form the basis for projections of anchovy and sardine under future climate change. They have shown that both bottom-up and top-down processes and species traits must be included. Two major challenges are the need to include climate change that is global in origin but regional in expression and the need to continue to improve our ability to characterize the relation of fish to the environment, particularly in terms of fish behavior.
CLIMATE CHANGE
GHG emissions cause warming of the earth, acidification of the ocean, and alteration of hydrology. These phenomena, in turn, affect anchovy and sardine in various ways, some known and others not. Projections for the future range from basic (including the poleward movement of populations with warming) to complex (based on the results of mechanistic models). Collectively, this spectrum of possible population responses begs for verification with observations.
Predicting Climate-Change Impacts on Anchovy and Sardine
The effects of future climate change (i.e., change attributed to increased GHG concentrations as well as changes in aerosols and land use) are necessarily based on assumptions regarding future emissions adding to current levels. Changes in atmospheric conditions associated with increased GHG concentrations are explored under scenarios of external forcing currently termed representative concentration pathways (RCPs) (IPCC 2013). The physics of global warming and the chemistry of ocean acidification are well understood and accepted. The subsequent effects on ocean dynamics and ecology, including anchovy and sardine, are less well understood.
Populations and stocks of anchovy and sardine inhabit regions of the global ocean, and therefore projections of change on the regional scale are needed. Climate change varies regionally (IPCC 2013). The finite but multiple potential effects of warming, acidification, and hydrology on small pelagic fish combined with the range of RCPs, regional variation, and uncertainty in future model projections lead to a large range of possible outcomes. The longer the projection is, the greater the uncertainties are. Long-term projections rely entirely on mechanistic models.
General circulation models are used to simulate the physical states of the atmosphere and ocean on the scale of seasons to millennia. They can offer probabilistic projections of climate over decadal to centennial timescales under assumed RCPs. Ensembles of general circulation model simulations are used to characterize variability within and between models. General circulation models are coupled with chemical and biological models to create earth system models. Models have been compared in the Coupled Model Intercomparison Project Phase 5 (IPCC 2013). During evaluations of the ability of models to make predictions of climate variability, testing may include withholding past data when fitting a model and then evaluating its ability to correctly reproduce the withheld data. Future predictions may be compared with observations to evaluate model skill over relatively short timescales. However, model assessment of climate-change projections is more challenging given the long prediction times and climate variability, which can inhibit direct comparison of model output with future observations and obscure detection of climate change in observational records.
Trust in model projections is essential for their use by decision makers. Trust is based, in part, on forecast skill as well as quantification of model uncertainty—i.e., how well different models agree and how likely long-term projections are to come to pass. Hence, skill evaluation must be a high priority. Weather forecasting benefits from the continuous assessment of forecast skill and subsequent improvement of forecast models. A similar protocol should exist for model forecasts of the effects of climate on anchovy and sardine. Interseasonal forecasts of Pacific sardine distribution in the Pacific Northwest are promising (Kaplan et al. 2016). Implicit in skill evaluation are observations adequate for this purpose. It is essential that time series of observations be sustained, curated, used, and expanded in places where fisheries-independent observations do not exist. Observation and skill assessment have been highlighted as key areas in climate-change research (IPCC 2013) and attribution science (Natl. Acad. Sci. Eng. Med. 2016).
Over the decadal to centennial timescales relevant for anthropogenic climate change, the ability to evaluate skill through comparisons between projections and observations is limited. Unlike forecasting efforts focused on changes at timescales of seasons to a year, it is impossible to quickly acquire the data necessary to assess model projections of changes that occur over many decades. Such projections rely on the ability of models to represent a wide range of mechanistic processes that can influence anchovy and sardine. Uncertainty of projections must be considered in a formal and systematic manner with an appreciation for differences among assumed RCPs, the formulations and resolutions of different models (including circulation, biogeochemical, fish, and higher-trophic-level models), natural variability in ecosystem conditions, and the role of mechanisms of change that may not be represented in current models (Cheung et al. 2016, Hawkins & Sutton 2009).
Observing
A common limitation for attribution is a sufficient observational time series (Natl. Acad. Sci. Eng. Med. 2016), which is necessary to characterize natural variability and hence know whether observations exceed natural bounds. Long time series of physical variables, such as sea level and temperature, are more common than those of fish abundance and distribution. Both fisheries-dependent and fisheries-independent data are useful. The world’s two longest time series of pelagic ecosystems are those of the California Cooperative Oceanic Fisheries Investigations (CalCOFI) and the Continuous Plankton Recorder (CPR), which have been running since 1949 and 1931, respectively. Time series are also needed to develop, execute, evaluate, and improve models.
General Projections
Potential processes that will influence the responses of anchovy and sardine to climate change include (a) geographic shifts in distribution (often poleward); (b) changes in phenology (seasonal, biological cycles), including the spring transition, spawning, migration, and predation mortality; (c) changes in the magnitude and sources of nutrients, including new nitrogen and nutrient ratios; (d) ocean acidification, including direct effects on fish and indirect effects on fish via the food web; (e) habitat compression caused by shoaling of waters high in CO2 and low in O2, altering trophic interactions; (f) changes in primary and secondary production and plankton species composition changes affecting trophic dynamics; (g) increased fishing mortality associated with changes in aquaculture and food security; and (h) acclimation and adaptation. One challenge is to be able to attribute observed changes to climate change rather than climate variability (Natl. Acad. Sci. Eng. Med. 2016). Time series of observations spanning multiple decades are often needed to do this, because climate variability occurs at multidecadal scales. For example, approximately 30–40 years of data are needed in most regions to differentiate between the influences of climate change and climate variability on phytoplankton (Henson et al. 2010). Because anchovy and sardine are strongly influenced by bottom-up processes, a similarly long (or longer) timescale is likely needed. Where attribution to climate change is not possible, consistency with it may be cited. Observed variability always includes natural variability (Natl. Acad. Sci. Eng. Med. 2016).
Change in a climate variable does not occur in isolation (Bopp et al. 2013). Warming alters stratification, habitable waters, and biological rates. Winds affect upwelling, flow, and mixing. Upwelled or mixed water is rich in nitrate but also enriched in CO2 and depleted in O2, with a low pH and carbonate saturation state and changes in other nutrients (Rykaczewski & Dunne 2010). Alteration of the hydrological cycle affects the amount, timing, and distribution of river discharge and thus nitrate and buoyancy fluxes to the ocean. Anchovy and sardine may respond differently to the same forces, reflecting their differing use of oceanic habitats and different diets.
Distribution shifts.
Warming is predicted to cause populations to move to higher latitudes and into deeper waters, although other movements may occur (Pinsky et al. 2013). Anchovy and sardine have moved northward into the North Sea and adjacent waters in the past few decades (Alheit et al. 2012, Beare et al. 2004, Montero-Serra et al. 2015, Petitgas et al. 2012). This shift is consistent with warming resulting from climate change. Although the occurrence of anchovy and sardine in these waters may not be unprecedented, it has been proposed as a forerunner of future changes (Enghoff et al. 2007).Phenology.
Changes in phenology are of particular concern because of their possible effects on trophic interactions, such as Cushing’s match-mismatch hypothesis (Durant et al. 2007). An analysis of the timing of the peak occurrence of anchovy and sardine larvae over six decades off California showed variations in phenology (Asch 2015). Although phenological changes for several species of larval fishes were consistent with climate change, variations in the phenology of anchovy and sardine were more consistent with an influence of climate variability.The migration phenology of forage fishes, including sardine, in the Columbia River plume was linked to seasonal changes in sea-surface temperature, salinity, and river flow and not linked to zooplankton abundance or upwelling (Kaltenberg & Benoit-Bird 2009). Although this study was too short to be directly informative about climate change, it indicates that changes in temperature and hydrology will influence sardine phenology under a warming climate. The phenology of river discharge, a source of new nitrogen, has changed in many regions and is projected to continue to change (Stewart et al. 2005). It also indicates that the phenology of sardine spawning and primary and secondary production may evolve differently under climate change, leading to potential seasonal mismatches, thus affecting mortality, growth rates, and recruitment.
Quality of source water.
An unexpected consequence of warming is a poleward shift in the subduction of source water upwelled in the California Current System (Rykaczewski & Dunne 2010). The increased transit time between sinking and upwelling of water masses leads to higher nitrate concentration and primary production in the habitat of anchovy and sardine. This mechanism also alters nutrient ratios in the upwelled water, which may affect the species composition of the plankton. Precipitation and river discharge will change, but not uniformly (IPCC 2013). Mean precipitation in midlatitude and subtropical regions is predicted to decrease in dry regions and increase in wet regions (IPCC 2013). Combined changes in climate and land use will cause the amount and nutrient content of river water to vary (IPCC 2013), with potentially greater effects on anchovy than on sardine.Ocean acidification.
The effects of ocean acidification on anchovy and sardine are perhaps the most challenging effects of climate change to describe. CO2 emitted by human activities accumulates in the ocean, decreasing the carbonate ion concentration (CO3−), pH, and saturation state of calcium carbonate (CaCO3) (Gattuso et al. 2015, IPCC 2013). The effects of these changes on anchovy and sardine may be direct, such as effects on olfaction (Munday et al. 2009) or balance and orientation (Checkley et al. 2009c), or indirect, via the food web (Bednaršek et al. 2014).Habitat compression.
Increased CO2 and decreased O2 move the aragonite compensation depth (Feely et al. 2008) and oxycline (Bertrand et al. 2011) upward, compressing the habitat for both competitors and predators of anchovy and sardine (Netburn & Koslow 2015).Plankton composition and production.
Primary production is projected to decrease at most subtropical and temperate latitudes where anchovy and sardine occur (Bopp et al. 2013). However, production in upwelling regions may increase and merit particular attention (Rykaczewski & Dunne 2010). Diatoms and other larger-sized phytoplankton are projected to experience more precipitous declines in abundance than other types of phytoplankton (Marinov et al. 2010). This has implications for anchovy and sardine, given their different feeding selectivities for plankton (van der Lingen et al. 2009). Climate-change effects on primary production are projected to become amplified at higher trophic levels (Stock et al. 2014). Gelatinous zooplankton may proliferate with warming, eutrophication, the depletion of predators and competitors by fishing, and increased substrate for reproduction, although there is little supporting evidence that ecosystems worldwide are becoming dominated by jellyfish (Condon et al. 2013). The increased abundance of whales and pinnipeds resulting from their protection (Estes et al. 2006) contributes to increased predation mortality of anchovy and sardine. Harmful algal blooms, which may be increasing because of climate change, killed sardine off South Africa while minimally affecting anchovy (van der Lingen et al. 2016). The finer filters of sardine are hypothesized to make them more vulnerable than anchovy to these blooms.Fishing.
Demand for small pelagic fish is likely to increase for use in aquaculture (Merino et al. 2010). Food security may be at risk owing to conflicts arising from climate change, drought, and changes in agricultural food production (Smith et al. 2011). Change in the availability of and demand for small pelagic fish will put stress on fisheries management. Distributional shifts caused by climate change and expansion or contraction of populations will require new transboundary, international efforts in management (Ishimura et al. 2013).Acclimation and adaptation.
Species may acclimate or adapt under climate change (Munday et al. 2013). Both experiments and observations are useful to assess these processes. Studies in locations where anchovy and sardine are exposed to conditions projected under climate change may be particularly valuable. Coastal Peru—the habitat for the anchoveta, which is the world’s largest single-species fishery—is such a place, with pCO2 ranging up to 1,500 ppm (Friederich et al. 2008). High levels of phenotypic plasticity may also reduce the ability of species to adapt to climate change via natural selection: Acclimation will initially lead to greater survival and resilience as change begins, but when conditions exceed the range to which a species has been previously exposed, acclimation may no longer be sufficient to promote resilience (Anderson et al. 2013).
SUMMARY POINTS
- Anchovy and sardine evolved over millions of years and share and differ in key traits.
- Anchovy and sardine are part of an assemblage of fish that eat plankton and are forage for predatory fish, squid, birds, and mammals. This assemblage may in places be dominated by migrating midwater fish rather than anchovy and sardine.
- Anchovy and sardine stocks have fluctuated, often asynchronously, on the scale of many decades for millennia. These fluctuations occurred both before and after the start of human fishing activity.
- Anchovy and sardine populations vary primarily in response to climate.
- Anchovy and sardine stocks can be classified by the source of new nitrogen supporting their food webs.
- Fishing may alter fluctuations in anchovy and sardine stocks, but it neither causes nor prevents these fluctuations.
- Climate change may affect anchovy and sardine populations through ocean warming, acidification, habitat compression, nutrient supply, phenology, and changes in the food web and demand for aquaculture.
- Realistic models of anchovy and sardine populations with climate change and fishing and at the appropriate scale and resolution are achievable.
- Trust in projections derives from continued assessment of forecast skill and model improvement.
FUTURE ISSUES
- Time series of observations of anchovy, sardine, and the environment are necessary to be able to attribute variations to climate change and for models.
- Mechanistic modeling of climate-change effects on anchovy and sardine should be at scales relevant to stocks, management, and policy.
- A focus is needed on bottom-up processes, particularly the supply of new nitrogen; top-down processes, including fishing; and species traits.
- Ocean acidification, acclimation, and adaptation merit particular attention and continued research, especially in regions of extreme natural variability in the environment, such as off Peru.
- Forecast skill must receive greater attention if model projections for use in decisions are to be trusted.
disclosure statement
The authors are not aware of any affiliations, memberships, funding, or financial holdings that might be perceived as affecting the objectivity of this review.
acknowledgments
D.M.C. was supported on sabbatical by the University of California, San Diego. R.G.A. was supported as a postdoctoral fellow at Princeton University by the Nippon Foundation and the University of British Columbia Nereus Program. R.R.R. was supported in part by National Science Foundation award #1434530. D.M.C. acknowledges the creative environment of Shaw Island. The authors thank Adi Khen for drawing Figure 3 and the Annual Review of Marine Science for inviting us to contribute this review.
literature cited
-
Agostini VN, Bakun A. 2002. ‘Ocean triads’ in the Mediterranean Sea: physical mechanisms potentially structuring reproductive habitat suitability (with example application to European anchovy, Engraulis encrasicolus). Fish. Oceanogr. 11:129–42
-
Alheit J, Bakun A. 2010. Population synchronies within and between ocean basins: apparent teleconnections and implications as to physical-biological linkage mechanisms. J. Mar. Syst. 79:267–85
-
Alheit J, Niquen M. 2004. Regime shifts in the Humboldt Current ecosystem. Prog. Oceanogr. 60:201–22
More AR articles citing this reference
-
Alheit J, Pohlmann T, Casini M, Greve W, Hinrichs R, et al. 2012. Climate variability drives anchovies and sardines into the North and Baltic Seas. Prog. Oceanogr. 96:128–39
-
Alheit J, Roy C, Kifani S. 2009. Decadal-scale variability in populations. See Checkley et al. 2009a, pp. 64–87
-
Anderson JJ, Gurarie E, Bracis C, Burke BJ, Laidre KL. 2013. Modeling climate change impacts on phenology and population dynamics of migratory marine species. Ecol. Model. 264:83–97
-
Asch RG. 2015. Climate change and decadal shifts in the phenology of larval fishes in the California Current ecosystem. PNAS 112:E4065–74
-
Atkinson A, Hill SL, Barange M, Pakhomov EA, Raubenheimer D, et al. 2014. Sardine cycles, krill declines, and locust plagues: revisiting ‘wasp-waist’ food webs. Trends Ecol. Evol. 29:309–16
-
Ayon P, Purca S, Guevara-Carrasco R. 2004. Zooplankton volume trends off Peru between 1964 and 2001. ICES J. Mar. Sci. 61:478–84
-
Ayon P, Swartzman G, Bertrand A, Gutierrez M, Bertrand S. 2008. Zooplankton and forage fish species off Peru: large-scale bottom-up forcing and local-scale depletion. Prog. Oceanogr. 79:208–14
-
Barange M, Bernal M, Cergole MC, Cubillos LA, Daskalov GM, et al. 2009a. Current trends in the assessment and management of stocks. See Checkley et al. 2009a, pp. 191–255
-
Barange M, Coetzee J, Takasuka A, Hill K, Gutierrez M, et al. 2009b. Habitat expansion and contraction in anchovy and sardine populations. Prog. Oceanogr. 83:251–60
-
Barange M, Merino G, Blanchard JL, Scholtens J, Harle J, et al. 2014. Impacts of climate change on marine ecosystem production in societies dependent on fisheries. Nat. Clim. Change 4:211–16
-
Baumgartner TR, Soutar A, Ferreira-Bartrina V. 1992. Reconstruction of the history of Pacific sardine and northern anchovy populations over the past two millennia from sediments of the Santa Barbara Basin, California. CalCOFI Rep. 33:24–40
-
Beare D, Burns F, Jones E, Peach K, Portilla E, et al. 2004. An increase in the abundance of anchovies and sardines in the north-western North Sea since 1995. Glob. Change Biol. 10:1209–13
-
Bednaršek N, Feely RA, Reum JCP, Peterson B, Menkel J, et al. 2014. Limacina helicina shell dissolution as an indicator of declining habitat suitability owing to ocean acidification in the California Current Ecosystem. Proc. R. Soc. B 281:20140123
More AR articles citing this reference
-
Bellier E, Planque B, Petitgas P. 2007. Historical fluctuations in spawning location of anchovy (Engraulis encrasicolus) and sardine (Sardina pilchardus) in the Bay of Biscay during 1967–73 and 2000–2004. Fish. Oceanogr. 16:1–15
-
Bertrand A, Chaigneau A, Peraltilla S, Ledesma J, Graco M, et al. 2011. Oxygen: a fundamental property regulating pelagic ecosystem structure in the coastal southeastern tropical Pacific. PLOS ONE 6:e29558
More AR articles citing this reference
-
Bertrand A, Segura M, Gutierrez M, Vasquez L. 2004. From small-scale habitat loopholes to decadal cycles: a habitat-based hypothesis explaining fluctuation in pelagic fish populations off Peru. Fish Fish. 5:296–316
-
Blaxter JHS, Hunter JR. 1982. The biology of the clupeoid fishes. In Advances in Marine Biology, Vol. 20, ed. JHS Blaxter, FS Russell, M Yonge, pp. 1–223. London: Academic
More AR articles citing this reference
-
Bopp L, Resplandy L, Orr JC, Doney SC, Dunne JP, et al. 2013. Multiple stressors of ocean ecosystems in the 21st century: projections with CMIP5 models. Biogeosciences 10:6225–45
More AR articles citing this reference
-
Bowen BW, Grant WS. 1997. Phylogeography of the sardines (Sardinops spp.): assessing biogeographic models and population histories in temperate upwelling zones. Evolution 51:1601–10
More AR articles citing this reference
-
Boyer DC, Boyer HJ, Fossen I, Kreiner A. 2001. Changes in abundance of the northern Benguela sardine stock during the decade 1990–2000, with comments on the relative importance of fishing and the environment. S. Afr. J. Mar. Sci. 23:67–84
More AR articles citing this reference
-
Boyer DC, Hampton I. 2001. An overview of the living marine resources of Namibia. S. Afr. J. Mar. Sci. 23:5–35
-
Brochier T, Echevin V, Tam J, Chaigneau A, Goubanova K, Bertrand A. 2013. Climate change scenarios experiments predict a future reduction in small pelagic fish recruitment in the Humboldt Current system. Glob. Change Biol. 19:1841–53
-
Carvalho FM, Castello JP. 2013. Argentine anchovy (Engraulis anchoita) stock identification and incipient exploitation in southern Brazil. Lat. Am. J. Aquat. Res. 41:820–27
-
Chavez FP, Ryan J, Lluch-Cota SE, Niquen M. 2003. From anchovies to sardines and back: multidecadal change in the Pacific Ocean. Science 299:217–21
More AR articles citing this reference
-
Checkley DM Jr., Alheit J, Oozeki Y, Roy C, eds. 2009a. Climate Change and Small Pelagic Fish. Cambridge, UK: Cambridge Univ. Press
-
Checkley DM Jr., Ayon P, Baumgartner TR, Bernal M, Coetzee JC, et al. 2009b. Habitats. See Checkley et al. 2009a, pp. 12–44
-
Checkley DM Jr., Dickson AG, Takahashi M, Radich JA, Eisenkolb N, Asch R. 2009c. Elevated CO2 enhances otolith growth in young fish. Science 324:1683
More AR articles citing this reference
-
Chelton DB, Schlax MG, Freilich MH, Milliff RF. 2004. Satellite measurements reveal persistent small-scale features in ocean winds. Science 303:978–83
More AR articles citing this reference
-
Cheung WWL, Frölicher TL, Asch RG, Jones MC, Pinsky ML, et al. 2016. Building confidence in projections of the responses of living marine resources to climate change. ICES J. Mar. Sci. 73:1283–96
-
Condon RH, Duarte CM, Pitt KA, Robinson KL, Lucas CH, et al. 2013. Recurrent jellyfish blooms are a consequence of global oscillations. PNAS 110:1000–5
-
Cury PM, Bakun A, Crawford RJM, Jarre A, Quinones RA, et al. 2000. Small pelagics in upwelling systems: patterns of interaction and structural changes in “wasp-waist” ecosystems. ICES J. Mar. Sci. 57:603–18
More AR articles citing this reference
-
Cury PM, Boyd IL, Bonhommeau S, Anker-Nilssen T, Crawford RJM, et al. 2011. Global seabird response to forage fish depletion: one-third for the birds. Science 334:1703–6
-
Daskalov GM, Boyer DC, Roux JP. 2003. Relating sardine Sardinops sagax abundance to environmental indices in northern Benguela. Prog. Oceanogr. 59:257–74
More AR articles citing this reference
-
Daskalov GM, Grishin AN, Rodionov S, Mihneva V. 2007. Trophic cascades triggered by overfishing reveal possible mechanisms of ecosystem regime shifts. PNAS 104:10518–23
More AR articles citing this reference
-
de Macedo-Soares LCP, Eiras Garcia CA, Freire AS, Muelbert JH. 2014. Large-scale ichthyoplankton and water mass distribution along the South Brazil Shelf. PLOS ONE 9:e91241
-
Demer DA, Zwolinski JP, Byers KA, Cutter GR, Renfree JS, et al. 2012. Prediction and confirmation of seasonal migration of Pacific sardine (Sardinops sagax) in the California Current Ecosystem. Fish. Bull. 110:52–70
-
Deyle ER, Fogarty M, Hsieh C-H, Kaufman L, MacCall AD, et al. 2013. Predicting climate effects on Pacific sardine. PNAS 110:6430–35
-
Dimmlich WF, Breed WG, Geddes M, Ward TM. 2004. Relative importance of gulf and shelf waters for spawning and recruitment of Australian anchovy, Engraulis australis, in South Australia. Fish. Oceanogr. 13:310–23
-
Dugdale RC, Goering JJ. 1967. Uptake of new and regenerated forms of nitrogen in primary productivity. Limnol. Oceanogr. 12:677–80
More AR articles citing this reference
-
Durant JM, Hjermann DO, Ottersen G, Stenseth NC. 2007. Climate and the match or mismatch between predator requirements and resource availability. Clim. Res. 33:271–83
-
Emmett RL, Brodeur RD, Miller TW, Pool SS, Krutzikowsky GK, et al. 2005. Pacific sardine (Sardinops sagax) abundance, distribution, and ecological relationships in the Pacific Northwest. CalCOFI Rep. 46:122–43
-
Enghoff IB, MacKenzie BR, Nielsen EE. 2007. The Danish fish fauna during the warm Atlantic period (ca. 7000–3900 BC): forerunner of future changes? Fish. Res. 87:167–80
-
Essington TE, Moriarty PE, Froehlich HE, Hodgson EE, Koehn LE, et al. 2015. Fishing amplifies forage fish population collapses. PNAS 112:6648–52
-
Estes J, Demaster D, Doak D, Williams T, Brownell R. 2006. Whales, Whaling and Ecosystems. Berkeley: Univ. Calif. Press
-
FAO (Food Agric. Organ. UN). 2014. The State of World Fisheries and Aquaculture. Rome: FAO
-
Feely RA, Sabine CL, Hernandez-Ayon JM, Ianson D, Hales B. 2008. Evidence for upwelling of corrosive “acidified” water onto the continental shelf. Science 320:1490–92
More AR articles citing this reference
-
Fiechter J, Rose KA, Curchitser EN, Hedstrom KS. 2015. The role of environmental controls in determining sardine and anchovy population cycles in the California Current: analysis of an end-to-end model. Prog. Oceanogr. 138:381–98
-
Field DB, Baumgartner TR, Ferreira V, Gutierrez D, Lozano-Montes H, et al. 2009. Variability from scales in marine sediments and other historical records. See Checkley et al. 2009a, pp. 45–63
-
Field JC, Francis RC, Aydin K. 2006. Top-down modeling and bottom-up dynamics: linking a fisheries-based ecosystem model with climate. Prog. Oceanogr. 68:238–70
More AR articles citing this reference
-
Finney BP, Alheit J, Emeis K-C, Field DB, Gutierrez D, Struck U. 2010. Paleoecological studies on variability in marine fish populations: a long-term perspective on the impacts of climatic change on marine ecosystems. J. Mar. Syst. 79:316–26
-
Friederich GE, Ledesma J, Ulloa O, Chavez FP. 2008. Air-sea carbon dioxide fluxes in the coastal southeastern tropical Pacific. Prog. Oceanogr. 79:156–66
-
Gattuso J-P, Magnan A, Billé R, Cheung WWL, Howes EL, et al. 2015. Contrasting futures for ocean and society from different anthropogenic CO2 emissions scenarios. Science 349:aac4722
More AR articles citing this reference
-
Glaser SM. 2011. Do albacore exert top-down pressure on northern anchovy? Estimating anchovy mortality as a result of predation by juvenile North Pacific albacore in the California Current system. Fish. Oceanogr. 20:242–57
-
Goericke R, Bograd SJ, Grundle DS. 2015. Denitrification and flushing of the Santa Barbara Basin bottom waters. Deep-Sea Res. II 112:53–60
-
Goldsworthy SD, Page B, Rogers PJ, Bulmand C, Wiebkin A, et al. 2013. Trophodynamics of the eastern Great Australian Bight ecosystem: ecological change associated with the growth of Australia’s largest fishery. Ecol. Model. 255:38–57
-
Grant WS, Bowen BW. 1998. Shallow population histories in deep evolutionary lineages of marine fishes: insights from sardines and anchovies and lessons for conservation. J. Hered. 89:415–26
-
Grant WS, Bowen BW. 2006. Living in a tilted world: climate change and geography limit speciation in Old World anchovies (Engraulis; Engraulidae). Biol. J. Linn. Soc. 88:673–89
-
Grant WS, Leslie RW, Bowen BW. 2005. Molecular genetic assessment of bipolarity in the anchovy genus Engraulis. J. Fish Biol. 67:1242–65
-
Guiñez M, Valdés J, Sifeddine A, Boussafir M, Dávila PM. 2014. Anchovy population and ocean-climatic fluctuations in the Humboldt Current System during the last 700 years and their implications. Palaeogeogr. Palaeoclim. Palaeoecol. 415:210–24
-
Guraslan C, Fach BA, Oguz T. 2014. Modeling the impact of climate variability on Black Sea anchovy recruitment and production. Fish. Oceanogr. 23:436–57
-
Gutierrez M, Swartzman G, Bertrand A, Bertrand S. 2007. Anchovy (Engraulis ringens) and sardine (Sardinops sagax) spatial dynamics and aggregation patterns in the Humboldt Current ecosystem, Peru, from 1983–2003. Fish. Oceanogr. 16:155–68
More AR articles citing this reference
-
Halouani G, Gascuel D, Hattab T, Lasram FBR, Coll M, et al. 2015. Fishing impact in Mediterranean ecosystems: an EcoTroph modeling approach. J. Mar. Syst. 150:22–33
-
Hawkins E, Sutton R. 2009. The potential to narrow uncertainty in regional climate predictions. Bull. Am. Meteorol. Soc. 90:1095–107
More AR articles citing this reference
-
Henson SA, Sarmiento JL, Dunne JP, Bopp L, Lima I, et al. 2010. Detection of anthropogenic climate change in satellite records of ocean chlorophyll and productivity. Biogeosciences 7:621–40
More AR articles citing this reference
-
Herrick SF Jr., Norton JG, Hannesson R, Sumaila UR, Ahmed M, Pena-Torres J. 2009. Global production and economics. See Checkley et al. 2009a, pp. 256–74
-
Holmgrenurba D, Baumgartner TR. 1993. A 250-year history of pelagic fish abundances from the anaerobic sediments of the central Gulf of California. CalCOFI Rep. 34:60–68
-
Hughes JM, Stewart J, Lyle JM, Suthers IM. 2014. Top-down pressure on small pelagic fish by eastern Australian salmon Arripis trutta; estimation of daily ration and annual prey consumption using multiple techniques. J. Exp. Mar. Biol. Ecol. 459:190–98
-
IPCC (Intergov. Panel Clim. Change). 2013. Summary for policymakers. In Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change, ed. TF Stocker, D Qin, G-K Plattner, M Tignor, SK Allen, et al., pp. 1–30. Cambridge, UK: Cambridge Univ. Press
-
Irigoien X, de Roos A. 2011. The role of intraguild predation in the population dynamics of small pelagic fish. Mar. Biol. 158:1683–90
-
Ishimura G, Herrick S, Sumaila UR. 2013. Stability of cooperative management of the Pacific sardine fishery under climate variability. Mar. Policy 39:333–40
-
Jacobson LD, Bograd SJ, Parrish RH, Mendelssohn R, Schwing FB. 2005. An ecosystem-based hypothesis for climatic effects on surplus production in California sardine (Sardinops sagax) and environmentally dependent surplus production models. Can. J. Fish. Aquat. Sci. 62:1782–96
-
Jahncke J, Checkley DM, Hunt GL. 2004. Trends in carbon flux to seabirds in the Peruvian upwelling system: effects of wind and fisheries on population regulation. Fish. Oceanogr. 13:208–23
More AR articles citing this reference
-
Jones WA. 2016. The Santa Barbara Basin fish assemblage in the last two millennia inferred from otoliths in sediment cores. PhD Thesis, Univ. Calif., San Diego
-
Kaltenberg AM, Benoit-Bird KJ. 2009. Diel behavior of sardine and anchovy schools in the California Current System. Mar. Ecol. Prog. Ser. 394:247–62
More AR articles citing this reference
-
Kamykowski D. 2012. 20th century variability of Atlantic meridional overturning circulation: planetary wave influences on world ocean surface phosphate utilization and synchrony of small pelagic fisheries. Deep-Sea Res. I 65:85–99
-
Kaplan IC, Williams GD, Bond NA, Hermann AJ, Siedlecki SA. 2016. Cloudy with a chance of sardines: forecasting sardine distributions using regional climate models. Fish. Oceanogr. 25:15–27
-
Kasapidis P. 2014. Phylogeography and population genetics. In Biology and Ecology of Sardines and Anchovies, ed. K Ganias, pp. 43–75. Boca Raton, FL: CRC Press
-
Kelleher K. 2005. Discards in the world’s marine fisheries: an update. Fish. Tech. Pap. 470, Food Agric. Organ. UN, Rome
-
Kirchner CH, Bartholomae CH, Kreiner A. 2009. Use of environmental parameters to explain the variability in spawner-recruitment relationships of Namibian sardine Sardinops sagax. Afr. J. Mar. Sci. 31:157–70
-
Koslow JA, Davison P, Lara-Lopez A, Ohman MD. 2014. Epipelagic and mesopelagic fishes in the southern California Current System: ecological interactions and oceanographic influences on their abundance. J. Mar. Syst. 138:20–28
-
Lecomte F, Grant WS, Dodson JJ, Rodriguez-Sanchez R, Bowen BW. 2004. Living with uncertainty: genetic imprints of climate shifts in East Pacific anchovy (Engraulis mordax) and sardine (Sardinops sagax). Mol. Ecol. 13:2169–82
-
Lindegren M, Checkley DM Jr., Rouyer T, MacCall AD, Stenseth NC. 2013. Climate, fishing, and fluctuations of sardine and anchovy in the California Current. PNAS 110:13672–77
-
Lloret J, Palomera I, Salat J, Sole I. 2004. Impact of freshwater input and wind on landings of anchovy (Engraulis encrasicolus) and sardine (Sardina pilchardus) in shelf waters surrounding the Ebre (Ebro) River delta (north-western Mediterranean). Fish. Oceanogr. 13:102–10
-
Lluch-Belda D, Lluch-Cota DB, Lluch-Cota SE. 2003. Baja California’s biological transition zones: refuges for the California sardine. J. Oceanogr. 59:503–13
-
Lynam CP, Gibbons MJ, Axelsen BE, Sparks CAJ, Coetzee J, et al. 2006. Jellyfish overtake fish in a heavily fished ecosystem. Curr. Biol. 16:R492–93
More AR articles citing this reference
-
MacCall AD. 1990. Dynamic Geography of Marine Fish Populations. Seattle: Wash. Sea Grant Program
-
MacCall AD. 2009. Mechanisms of low-frequency fluctuations in sardine and anchovy populations. See Checkley et al. 2009a, pp. 285–99
-
MacCall AD, Sydeman WJ, Davison PC, Thayer JA. 2016. Recent collapse of northern anchovy biomass off California. Fish. Res. 175:87–94
-
MacDonald GM, Case RA. 2005. Variations in the Pacific Decadal Oscillation over the past millennium. Geophys. Res. Lett. 32:L08703
-
Marinov I, Doney SC, Lima ID. 2010. Response of ocean phytoplankton community structure to climate change over the 21st century: partitioning the effects of nutrients, temperature and light. Biogeosciences 7:3941–59
-
Merino G, Barange M, Mullon C. 2010. Climate variability and change scenarios for a marine commodity: modelling small pelagic fish, fisheries and fishmeal in a globalized market. J. Mar. Syst. 81:196–205
-
Minobe S. 1997. A 50–70 year climatic oscillation over the North Pacific and North America. Geophys. Res. Lett. 24:683–86
More AR articles citing this reference
-
Montero-Serra I, Edwards M, Genner MJ. 2015. Warming shelf seas drive the subtropicalization of European pelagic fish communities. Glob. Change Biol. 21:144–53
-
Munday PL, Dixson DL, Donelson JM, Jones GP, Pratchett MS, et al. 2009. Ocean acidification impairs olfactory discrimination and homing ability of a marine fish. PNAS 106:1848–52
More AR articles citing this reference
-
Munday PL, Warner RR, Monro K, Pandolfi JM, Marshall DJ. 2013. Predicting evolutionary responses to climate change in the sea. Ecol. Lett. 16:1488–500
More AR articles citing this reference
-
Murphy GI. 1968. Pattern in life history and the environment. Am. Nat. 102:391–403
More AR articles citing this reference
-
Natl. Acad. Sci. Eng. Med. 2016. Attribution of Extreme Weather Events in the Context of Climate Change. Washington, DC: Natl. Acad. Press
-
Netburn AN, Koslow JA. 2015. Dissolved oxygen as a constraint on daytime deep scattering layer depth in the southern California current ecosystem. Deep-Sea Res. I 104:149–58
-
Nishikawa H, Yasuda I. 2008. Japanese sardine (Sardinops melanostictus) mortality in relation to the winter mixed layer depth in the Kuroshio Extension region. Fish. Oceanogr. 17:411–20
-
Nishikawa H, Yasuda I. 2011. Long-term variability of winter mixed layer depth and temperature along the Kuroshio jet in a high-resolution ocean general circulation model. J. Oceanogr. 67:503–18
-
O’Donoghue SH, Whittington PA, Dyer BM, Peddemors VM. 2010. Abundance and distribution of avian and marine mammal predators of sardine observed during the 2005 KwaZulu-Natal sardine run survey. Afr. J. Mar. Sci. 32:361–74
-
Oguz T, Fach B, Salihoglu B. 2008. Invasion dynamics of the alien ctenophore Mnemiopsis leidyi and its impact on anchovy collapse in the Black Sea. J. Plankton Res. 30:1385–97
More AR articles citing this reference
-
Okunishi T, Ito S, Hashioka T, Sakamoto TT, Yoshie N, et al. 2012. Impacts of climate change on growth, migration and recruitment success of Japanese sardine (Sardinops melanostictus) in the western North Pacific. Clim. Change 115:485–503
-
Oozeki Y, Takasuka A, Kubota H, Barange M. 2007. Characterizing spawning habitats of Japanese sardine (Sardinops melanostictus), Japanese anchovy (Engraulis japonicus), and Pacific round herring (Etrumeus teres) in the northwestern Pacific. CalCOFI Rep. 48:191–203
-
Ospina-Alvarez A, Catalán IA, Bernal M, Roos D, Palomera I. 2015. From egg production to recruits: connectivity and inter-annual variability in the recruitment patterns of European anchovy in the northwestern Mediterranean. Prog. Oceanogr. 138:431–47
-
Parker RWR, Tyedmers PH. 2015. Fuel consumption of global fishing fleets: current understanding and knowledge gaps. Fish Fish. 16:684–96
-
Parrish RH, Serra R, Grant WS. 1989. The monotypic sardines, Sardina and Sardinops: their taxonomy, distribution, stock structure and zoogeography. Can. J. Fish. Aquat. Sci. 46:2019–36
-
Pauly D, Christensen V, Walters C. 2000. Ecopath, Ecosim, and Ecospace as tools for evaluating ecosystem impact of fisheries. ICES J. Mar. Sci. 57:697–706
More AR articles citing this reference
-
Pauly D, Zeller D. 2016. Catch reconstructions reveal that global marine fisheries catches are higher than reported and declining. Nat. Commun. 7:10244
More AR articles citing this reference
-
Petitgas P, Alheit J, Peck MA, Raab K, Irigoien X, et al. 2012. Anchovy population expansion in the North Sea. Mar. Ecol. Prog. Ser. 444:1–13
-
Pinsky ML, Byler D. 2015. Fishing, fast growth and climate variability increase the risk of collapse. Proc. R. Soc. B 282:20151053
-
Pinsky ML, Worm B, Fogarty MJ, Sarmiento JL, Levin SA. 2013. Marine taxa track local climate velocities. Science 341:1239–42
-
Piroddi C, Coll M, Steenbeek J, Moy DM, Christensen V. 2015. Modelling the Mediterranean marine ecosystem as a whole: addressing the challenge of complexity. Mar. Ecol. Prog. Ser. 533:47–65
-
Politikos D, Somarakis S, Tsiaras KP, Giannoulaki M, Petihakis G, et al. 2015. Simulating anchovy’s full life cycle in the northern Aegean Sea (eastern Mediterranean): a coupled hydro-biogeochemical-IBM model. Prog. Oceanogr. 138:399–416
-
Quinn TJ, Deriso RB. 1999. Quantitative Fisheries Dynamics. Oxford, UK: Oxford Univ. Press
-
Robinson KR, Ruzicka JJ, Decker MB, Brodeur RD, Hernandez FJ, et al. 2014. Jellyfish, forage fish, and the world’s major fisheries. Oceanography 27(4):104–15
-
Rose KA, Fiechter J, Curchitser EN, Hedstrom K, Bernal M, et al. 2015. Demonstration of a fully-coupled end-to-end model for small pelagic fish using sardine and anchovy in the California Current. Prog. Oceanogr. 138:348–80
-
Roux J-P, van der Lingen CD, Gibbons MJ, Moroff NE, Shannon LJ, et al. 2013. Jellyfication of marine ecosystems as a likely consequence of overfishing small pelagic fishes: lessons from the Benguela. Bull. Mar. Sci. 89:249–84
-
Ruzicka JJ, Brodeur RD, Emmett RL, Steele JH, Zamon JE, et al. 2012. Interannual variability in the Northern California Current food web structure: changes in energy flow pathways and the role of forage fish, euphausiids, and jellyfish. Prog. Oceanogr. 102:19–41
More AR articles citing this reference
-
Rykaczewski RR, Checkley DM. 2008. Influence of ocean winds on the pelagic ecosystem in upwelling regions. PNAS 105:1965–70
More AR articles citing this reference
-
Rykaczewski RR, Dunne JP. 2010. Enhanced nutrient supply to the California Current Ecosystem with global warming and increased stratification in an earth system model. Geophys. Res. Lett. 37:L21606
More AR articles citing this reference
-
Ryther JH. 1969. Photosynthesis and fish production in the sea. Science 166:72–76
More AR articles citing this reference
-
Sanchez RP, Deciechomski JD. 1995. Spawning and nursery grounds of pelagic fish species in the sea-shelf off Argentina and adjacent areas. Sci. Mar. 59:455–78
-
Sea Around Us. 2016. Sea Around Us: fisheries, ecosystems and biodiversity. Accessed Apr. 5, 2016. Univ. B.C., Vancouver, Can. http://www.seaaroundus.org
-
Shackleton LY. 1987. A comparative-study of fossil fish scales from three upwelling regions. S. Afr. J. Mar. Sci. 5:79–84
More AR articles citing this reference
-
Shannon L, Coll M, Neira S, Cury PM, Roux J-P. 2009. Impacts of fishing and climate change explored using trophic models. See Checkley et al. 2009a, pp. 158–90
-
Smith ADM, Brown CJ, Bulman CM, Fulton EA, Johnson P, et al. 2011. Impacts of fishing low-trophic level species on marine ecosystems. Science 333:1147–50
More AR articles citing this reference
-
Smith MD, Fulton EA, Day RW. 2015. Using an Atlantis model of the southern Benguela to explore the response of ecosystem indicators for fisheries management. Environ. Model. Softw. 69:23–41
-
Smith PE. 2005. A history of proposals for subpopulation structure in the Pacific sardine (Sardinops sagax) population of western North America. CalCOFI Rep. 46:75–82
-
Soutar A, Isaacs JD. 1969. History of fish populations inferred from fish scales in anaerobic sediments off California. CalCOFI Rep. 13:63–70
-
Steele JH, Ruzicka JJ. 2011. Constructing end-to-end models using ECOPATH data. J. Mar. Syst. 87:227–38
More AR articles citing this reference
-
Stewart IT, Cayan DR, Dettinger MD. 2005. Changes toward earlier streamflow timing across western North America. J. Clim. 18:1136–55
More AR articles citing this reference
-
Stock CA, Dunne JP, John JG. 2014. Drivers of trophic amplification of ocean productivity trends in a changing climate. Biogeosciences 11:7125–35
-
Swartzman G, Bertrand A, Gutierrez M, Bertrand S, Vasquez L. 2008. The relationship of anchovy and sardine to water masses in the Peruvian Humboldt Current System from 1983 to 2005. Prog. Oceanogr. 79:228–37
-
Takasuka A, Oozeki Y, Aoki I. 2007. Optimal growth temperature hypothesis: Why do anchovy flourish and sardine collapse or vice versa under the same ocean regime? Can. J. Fish. Aquat. Sci. 64:768–76
-
Takasuka A, Oozeki Y, Kimura R, Kubota H, Aoki I. 2004. Growth-selective predation hypothesis revisited for larval anchovy in offshore waters: cannibalism by juveniles versus predation by skipjack tunas. Mar. Ecol. Prog. Ser. 278:297–302
-
Travers-Trolet M, Shin YJ, Field JG. 2014. An end-to-end coupled model ROMS-N2P2Z2D2-OSMOSE of the southern Benguela foodweb: parameterisation, calibration and pattern-oriented validation. Afr. J. Mar. Sci. 36:11–29
-
Tsai CF, Chen PY, Chen CP, Lee MA, Shiah GY, Lee KT. 1997. Fluctuation in abundance of larval anchovy and environmental conditions in coastal waters off south-western Taiwan as associated with the El Niño Southern Oscillation. Fish. Oceanogr. 6:238–49
-
Tu C-Y, Tseng Y-H, Chiu T-S, Shen M-L, Hsieh C-H. 2012. Using coupled fish behavior–hydrodynamic model to investigate spawning migration of Japanese anchovy, Engraulis japonicus, from the East China Sea to Taiwan. Fish. Oceanogr. 21:255–68
-
Valdes J, Ortlieb L, Gutierrez D, Marinovic L, Vargas G, Sifeddine A. 2008. 250 years of sardine and anchovy scale deposition record in Mejillones Bay, northern Chile. Prog. Oceanogr. 79:198–207
-
van der Lingen CD, Bertrand A, Bode A, Brodeur R, Cubillos LA, et al. 2009. Trophic dynamics. See Checkley et al. 2009a, pp. 112–57
-
van der Lingen CD, Coetzee JC, Hutchings L. 2010. Overview of the KwaZulu-Natal sardine run. Afr. J. Mar. Sci. 32:271–77
-
van der Lingen CD, Hutchings L, Lamont T, Pitcher GC. 2016. Climate change, dinoflagellate blooms and sardine in the southern Benguela Current Large Marine Ecosystem. Environ. Dev. 17(Suppl. 1):230–43
-
Ward TM, Hoedt F, McLeay L, Dimmlich WF, Kinloch M, et al. 2001. Effects of the 1995 and 1998 mass mortality events on the spawning biomass of sardine, Sardinops sagax, in South Australian waters. ICES J. Mar. Sci. 58:865–75
-
Watanabe Y. 2009. Recruitment variability of small pelagic fish populations in the Kuroshio-Oyashio transition region of the western North Pacific. J. Northwest Atl. Fish. Sci. 41:197–204
-
Watanabe Y, Zenitani H, Kimura R. 1995. Population decline of the Japanese sardine Sardinops melanostictus owing to recruitment failures. Can. J. Fish. Aquat. Sci. 52:1609–16
-
Xu Y, Rose KA, Chai F, Chavez FP, Ayon P. 2015. Does spatial variation in environmental conditions affect recruitment? A study using a 3-D model of Peruvian anchovy. Prog. Oceanogr. 138:417–30
-
Yasuda I. 2003. Hydrographic structure and variability in the Kuroshio-Oyashio Transition Area. J. Oceanogr. 59:389–402
-
Zarauz L, Irigoien X, Fernandes JA. 2009. Changes in plankton size structure and composition, during the generation of a phytoplankton bloom, in the central Cantabrian sea. J. Plankton Res. 31:193–207
-
Zarraonaindia I, Iriondo M, Albaina A, Pardo MA, Manzano C, et al. 2012. Multiple SNP markers reveal fine-scale population and deep phylogeographic structure in European anchovy (Engraulis encrasicolus L.). PLOS ONE 7:e42201
RELATED RESOURCES
CalCOFI (Calif. Coop. Ocean. Fish. Investig.): http://www.calcofi.org
Ganias K, ed. 2014. Biology and Ecology of Sardines and Anchovies. Boca Raton, FL: CRC Press
SAHFOS (Sir Alistair Hardy Found. Ocean Sci.) Continuous Plankton Recorder: http://www.sahfos.ac.uk